
The domain within your query sequence starts at position 763 and ends at position 958; the E-value for the CYCc domain shown below is 2.68e-107.
All catalytic sites are present in this domain. Check the literature (PubMed 97222132 ) for details.
AERDRADHLNFMLLPRLVVKSLKEKGIVEPELYEEVTIYFSDIVGFTTICKYSTPMEVVD MLNDIYKSFDQIVDHHDVYKVETIGDAYVVASGLPMRNGNRHAVDISKMALDILSFIGTF ELEHLPGLPVWIRIGVHSGPCAAGVVGIKMPRYCLFGDTVNTASRMESTGLPLRIHMSSS TITILKRTDCQFLYEV
CYCcAdenylyl- / guanylyl cyclase, catalytic domain |
|---|
| SMART accession number: | SM00044 |
|---|---|
| Description: | Present in two copies in mammalian adenylyl cyclases. Eubacterial homologues are known. Two residues (Asn, Arg) are thought to be involved in catalysis. These cyclases have important roles in a diverse range of cellular processes. |
| Interpro abstract (IPR001054): | Guanylate cyclases ( EC 4.6.1.2 ) catalyse the formation of cyclic GMP (cGMP) from GTP. cGMP acts as an intracellular messenger, activating cGMP-dependent kinases and regulating cGMP-sensitive ion channels. The role of cGMP as a second messenger in vascular smooth muscle relaxation and retinal photo-transduction is well established. Guanylate cyclase is found both in the soluble and particulate fractions of eukaryotic cells. The soluble and plasma membrane-bound forms differ in structure, regulation and other properties [ (PUBMED:1349465) (PUBMED:1356629) (PUBMED:1680765) (PUBMED:1982420) ]. Most currently known plasma membrane-bound forms are receptors for small polypeptides. The soluble forms of guanylate cyclase are cytoplasmic heterodimers having alpha and beta subunits. In all characterised eukaryote guanylyl- and adenylyl cyclases, cyclic nucleotide synthesis is carried out by the conserved class III cyclase domain. |
| GO process: | cyclic nucleotide biosynthetic process (GO:0009190), intracellular signal transduction (GO:0035556) |
| Family alignment: |
There are 67904 CYCc domains in 62634 proteins in SMART's nrdb database.
Click on the following links for more information.
- Evolution (species in which this domain is found)
-
Taxonomic distribution of proteins containing CYCc domain.
This tree includes only several representative species. The complete taxonomic breakdown of all proteins with CYCc domain is also avaliable.
Click on the protein counts, or double click on taxonomic names to display all proteins containing CYCc domain in the selected taxonomic class.
- Cellular role (predicted cellular role)
-
Binding / catalysis: cAMP-synthesis, cGMP-synthesis
- Literature (relevant references for this domain)
-
Primary literature is listed below; Automatically-derived, secondary literature is also avaliable.
- Hurley JH
- Structure, mechanism, and regulation of mammalian adenylyl cyclase.
- J Biol Chem. 1999; 274: 7599-602
- Tucker CL, Hurley JH, Miller TR, Hurley JB
- Two amino acid substitutions convert a guanylyl cyclase, RetGC-1, into an adenylyl cyclase.
- Proc Natl Acad Sci U S A. 1998; 95: 5993-7
- Display abstract
Guanylyl cyclases (GCs) and adenylyl cyclases (ACs) have fundamental roles in a wide range of cellular processes. Whereas GCs use GTP as a substrate to form cGMP, ACs catalyze the analogous conversion of ATP to cAMP. Previously, a model based on the structure of adenylate cyclase was used to predict the structure of the nucleotide-binding pocket of a membrane guanylyl cyclase, RetGC-1. Based on this model, we replaced specific amino acids in the guanine-binding pocket of GC with their counterparts from AC. A change of two amino acids, E925K together with C995D, is sufficient to completely alter the nucleotide specificity from GTP to ATP. These experiments strongly validate the AC-derived RetGC-1 structural model and functionally confirm the role of these residues in nucleotide discrimination.
- Artymiuk PJ, Poirrette AR, Rice DW, Willett P
- A polymerase I palm in adenylyl cyclase?
- Nature. 1997; 388: 33-4
- Hobbs AJ
- Soluble guanylate cyclase: the forgotten sibling.
- Trends Pharmacol Sci. 1997; 18: 484-91
- Display abstract
Despite widespread distribution in most mammalian cells, the role of soluble guanylate cyclase has, until recently, been poorly defined, especially when compared with its more illustrious sibling, adenylate cyclase. In this review Adrian Hobbs outlines some of the reasons why the soluble guanylate cyclase-cGMP pathway has remained outside the signalling spotlight for much of the past 30 years. He goes on to describe how new molecular biological and biochemical approaches have facilitated a characterization of soluble guanylate cyclase and how this enzyme has acquired a profound physiological significance, and much research attention, as the intracellular 'receptor' for nitric oxide.
- Liu Y, Ruoho AE, Rao VD, Hurley JH
- Catalytic mechanism of the adenylyl and guanylyl cyclases: modeling and mutational analysis.
- Proc Natl Acad Sci U S A. 1997; 94: 13414-9
- Display abstract
The adenylyl and guanylyl cyclases catalyze the formation of 3', 5'-cyclic adenosine or guanosine monophosphate from the corresponding nucleoside 5'-triphosphate. The guanylyl cyclases, the mammalian adenylyl cyclases, and their microbial homologues function as pairs of homologous catalytic domains. The crystal structure of the rat type II adenylyl cyclase C2 catalytic domain was used to model by homology a mammalian adenylyl cyclase C1-C2 domain pair, a homodimeric adenylyl cyclase of Dictyostelium discoideum, a heterodimeric soluble guanylyl cyclase, and a homodimeric membrane guanylyl cyclase. Mg2+ATP or Mg2+GTP were docked into the active sites based on known stereochemical constraints on their conformation. The models are consistent with the activities of seven active-site mutants. Asp-310 and Glu-432 of type I adenylyl cyclase coordinate a Mg2+ ion. The D310S and D310A mutants have 10-fold reduced Vmax and altered [Mg2+] dependence. The NTP purine moieties bind in mostly hydrophobic pockets. Specificity is conferred by a Lys and an Asp in adenylyl cyclase, and a Glu, an Arg, and a Cys in guanylyl cyclase. The models predict that an Asp from one domain is a general base in the reaction, and that the transition state is stabilized by a conserved Asn-Arg pair on the other domain.
- vanHaastert PJ, Kuwayama H
- cGMP as second messenger during Dictyostelium chemotaxis.
- FEBS Lett. 1997; 410: 25-8
- Display abstract
The chemoattractant cAMP induces directed cell locomotion in Dictyostelium cells. Several second messenger pathways are activated upon binding of cAMP to G-protein-coupled receptors, including adenylyl cyclase, guanylyl cyclase, phospholipase C, and the opening of plasma membrane Ca2+ channels. These second messenger responses are unaltered in many chemotactic mutants, except for the cGMP response. Activation of guanylyl cyclase depends on G-proteins and is regulated by a cGMP-binding protein in a complex manner. This cGMP-binding protein also mediates intracellular functions of cGMP to activate a PKC-related kinase that phosphorylates myosin II heavy chain, thereby allowing myosin filaments to rearrange during cell movement.
- Wedel BJ, Garbers DL
- New insights on the functions of the guanylyl cyclase receptors.
- FEBS Lett. 1997; 410: 29-33
- Display abstract
The discovery of at least 29 genes encoding putative guanylyl cyclases in Caenorhabditis elegans has raised the question as to whether there are numerous receptors yet to be discovered in the mammal. The nematode, however, not only seems ideal to study guanylyl cyclase receptor localization and function, given the large variety of isoforms, but also leads to possible identification of ligands for orphan guanylyl cyclases by the use of genetic and behavioral assays. A recent powerful approach to describe the function of different guanylyl cyclase isoforms in mammals has been the disruption of the corresponding genes in the mouse. A salt resistant elevation of blood pressure, which corresponds to the phenotype of 50% of all human patients with essential hypertension, is observed in mice lacking the GC-A-receptor. Mice missing the GC-C receptor have been shown to be resistant to STa, an E. coli heat-stable enterotoxin, which is largely responsible for travellers diarrhea in adults and mortality due to diarrhea in infants.
- Yan SZ, Huang ZH, Shaw RS, Tang WJ
- The conserved asparagine and arginine are essential for catalysis of mammalian adenylyl cyclase.
- J Biol Chem. 1997; 272: 12342-9
- Display abstract
Mammalian adenylyl cyclases have two homologous cytoplasmic domains (C1 and C2), and both domains are required for the high enzymatic activity. Mutational and genetic analyses of type I and soluble adenylyl cyclases suggest that the C2 domain is catalytically active and the C1 domain is not; the role of the C1 domain is to promote the catalytic activity of the C2 domain. Two amino acid residues, Asn-1025 and Arg-1029 of type II adenylyl cyclase, are conserved among the C2 domains, but not among the C1 domains, of adenylyl cyclases with 12 putative transmembrane helices. Mutations at each amino acid residue alone result in a 30-100-fold reduction in Kcat of adenylyl cyclase. However, the same mutations do not affect the Km for ATP, the half-maximal concentration (EC50) for the C2 domain of type II adenylyl cyclase to associate with the C1 domain of type I adenylyl cyclase and achieve maximal enzyme activity, or the EC50 for forskolin to maximally activate enzyme activity with or without Gsalpha. This indicates that the mutations at these two residues do not cause gross structural alteration. Thus, these two conserved amino acid residues appear to be crucial for catalysis, and their absence from the C1 domains may account for its lack of catalytic activity. Mutations at both amino acid residues together result in a 3,000-fold reduction in Kcat of adenylyl cyclase, suggesting that these two residues have additive effects in catalysis. A second site suppressor of the Asn-1025 to Ser mutant protein has been isolated. This suppressor has 17-fold higher activity than the mutant and has a Pro-1015 to Ser mutation.
- Zhang G, Liu Y, Ruoho AE, Hurley JH
- Structure of the adenylyl cyclase catalytic core.
- Nature. 1997; 386: 247-53
- Display abstract
Mammalian adenylyl cyclases contain two conserved regions, C1 and C2, which are responsible for forskolin- and G-protein-stimulated catalysis. The structure of the C2 catalytic region of type II rat adenylyl cyclase has an alpha/beta class fold in a wreath-like dimer, which has a central cleft. Two forskolin molecules bind in hydrophobic pockets at the ends of cleft. The central part of the cleft is lined by charged residues implicated in ATP binding. Forskolin appears to activate adenylyl cyclase by promoting the assembly of the active dimer and by direct interaction within the catalytic cleft. Other adenylyl cyclase regulators act at the dimer interface or on a flexible C-terminal region.
- Pitt GS, Milona N, Borleis J, Lin KC, Reed RR, Devreotes PN
- Structurally distinct and stage-specific adenylyl cyclase genes play different roles in Dictyostelium development.
- Cell. 1992; 69: 305-15
- Display abstract
We have isolated two adenylyl cyclase genes, designated ACA and ACG, from Dictyostelium. The proposed structure for ACA resembles that proposed for mammalian adenylyl cyclases: two large hydrophilic domains and two sets of six transmembrane spans. ACG has a novel structure, reminiscent of the membrane-bound guanylyl cyclases. An aca- mutant, created by gene disruption, has little detectable adenylyl cyclase activity and fails to aggregate, demonstrating that cAMP is required for cell-cell communication. cAMP is not required for motility, chemotaxis, growth, and cell division, which are unaffected. Constitutive expression in aca- cells of either ACA or ACG, which is normally expressed only during germination, restores aggregation and the ability to complete the developmental program. ACA expression restores receptor and guanine nucleotide-regulated adenylyl cyclase activity, while activity in cells expressing ACG is insensitive to these regulators. Although they lack ACA, which has a transporter-like structure, the cells expressing ACG secrete cAMP constitutively.
- Tang WJ, Gilman AG
- Adenylyl cyclases.
- Cell. 1992; 70: 869-72
- Disease (disease genes where sequence variants are found in this domain)
-
SwissProt sequences and OMIM curated human diseases associated with missense mutations within the CYCc domain.
Protein Disease Retinal guanylyl cyclase 1 (Q02846) (SMART) OMIM:601777: Cone dystrophy, progressive
OMIM:600179: Leber congenital amaurosis, type I
OMIM:204000: Cone-rod dystrophy 6
OMIM:601777: - Metabolism (metabolic pathways involving proteins which contain this domain)
-
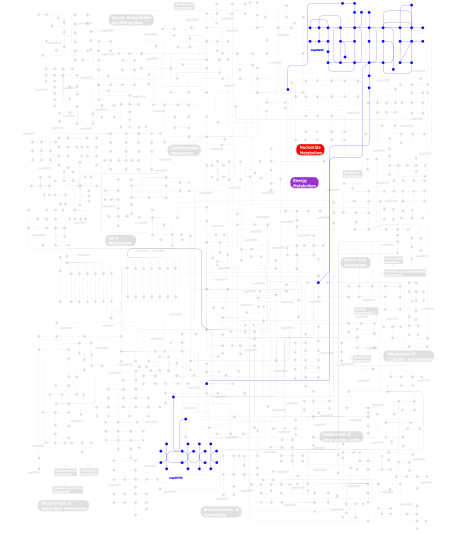
Click the image to view the interactive version of the map in iPath% proteins involved KEGG pathway ID Description 47.17  map00230
map00230Purine metabolism 15.66 map04540 Gap junction 9.49 map04730 Long-term depression 6.36 map04914 Progesterone-mediated oocyte maturation 6.16 map04912 GnRH signaling pathway 6.16 map04916 Melanogenesis 4.95 map04020 Calcium signaling pathway 1.82 map04742 Taste transduction 1.41 map04720 Long-term potentiation 0.71 map04740 Olfactory transduction 0.10 map00190Oxidative phosphorylation This information is based on mapping of SMART genomic protein database to KEGG orthologous groups. Percentage points are related to the number of proteins with CYCc domain which could be assigned to a KEGG orthologous group, and not all proteins containing CYCc domain. Please note that proteins can be included in multiple pathways, ie. the numbers above will not always add up to 100%.
- Structure (3D structures containing this domain)
3D Structures of CYCc domains in PDB
PDB code Main view Title 1ab8 
RAT TYPE II ADENYLYL CYCLASE C2 DOMAIN/FORSKOLIN COMPLEX 1azs 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE 1cjk 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE: COMPLEX WITH ADENOSINE 5'-(ALPHA THIO)-TRIPHOSPHATE (RP), MG, AND MN 1cjt 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE: COMPLEX WITH BETA-L-2',3'-DIDEOXYATP, MN, AND MG 1cju 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE: COMPLEX WITH BETA-L-2',3'-DIDEOXYATP AND MG 1cjv 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE: COMPLEX WITH BETA-L-2',3'-DIDEOXYATP, MG, AND ZN 1cs4 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE: COMPLEX WITH 2'-DEOXY-ADENOSINE 3'-MONOPHOSPHATE, PYROPHOSPHATE AND MG 1cul 
COMPLEX OF GS-ALPHA WITH THE CATALYTIC DOMAINS OF MAMMALIAN ADENYLYL CYCLASE: COMPLEX WITH 2',5'-DIDEOXY-ADENOSINE 3'-TRIPHOSPHATE AND MG 1fx2 
STRUCTURAL ANALYSIS OF ADENYLATE CYCLASES FROM TRYPANOSOMA BRUCEI IN THEIR MONOMERIC STATE 1fx4 
STRUCTURE ANALYSIS OF ADENYLATE CYCLASES FROM TRYPANOSOMA BRUCEI IN THEIR MONOMERIC STATE 1tl7 
Complex Of Gs- With The Catalytic Domains Of Mammalian Adenylyl Cyclase: Complex With 2'(3')-O-(N-methylanthraniloyl)-guanosine 5'-triphosphate and Mn 1u0h 
STRUCTURAL BASIS FOR THE INHIBITION OF MAMMALIAN ADENYLYL CYCLASE BY MANT-GTP 1wc0 
Soluble adenylyl cyclase CyaC from S. platensis in complex with alpha, beta-methylene-ATP 1wc1 
Soluble adenylyl cyclase CyaC from S. platensis in complex with Rp- ATPalphaS 1wc3 
Soluble adenylyl cyclase CyaC from S. platensis in complex with alpha, beta-methylene-ATP 1wc4 
Soluble adenylyl cyclase CyaC from S. platensis in complex with alpha, beta-methylene-ATP 1wc5 
Soluble adenylyl cyclase CyaC from S. platensis in complex with alpha, beta-methylene-ATP 1wc6 
Soluble adenylyl cyclase CyaC from S. platensis in complex with Rp- ATPalphaS in presence of bicarbonate 1y10 
Mycobacterial adenylyl cyclase Rv1264, holoenzyme, inhibited state 1y11 
Mycobacterial adenylyl cyclase Rv1264, holoenzyme, active state 1ybt 
MYCOBACTERIUM TUBERCULOSIS ADENYLYL CYCLASE, RV1900C CHD 1ybu 
Mycobacterium tuberculosis adenylyl cyclase Rv1900c CHD, in complex with a substrate analog. 1yk9 
Crystal structure of a mutant form of the mycobacterial adenylyl cyclase Rv1625c 2bw7 
A novel mechanism for adenylyl cyclase inhibition from the crystal structure of its complex with catechol estrogen 2gvd 
Complex Of Gs- With The Catalytic Domains Of Mammalian Adenylyl Cyclase: Complex With TNP-ATP and Mn 2gvz 
Crystal Structure of Complex of Gs- with The Catalytic Domains of Mammalian Adenylyl Cyclase: Complex with MANT-ATP and Mn 2w01 
Crystal structure of the guanylyl cyclase Cya2 2wz1 
STRUCTURE OF THE CATALYTIC DOMAIN OF HUMAN SOLUBLE GUANYLATE CYCLASE 1 BETA 3. 3c14 
Complex of GS-Alpha with the Catalytic Domains of Mammalian Adenylyl Cyclase: Complex with Pyrophosphate and Ca 3c15 
Complex of GS-Alpha with the Catalytic Domains of Mammalian Adenylyl Cyclase: Complex with Pyrophosphate and Mg 3c16 
Complex of GS-Alpha with the Catalytic Domains of Mammalian Adenylyl Cyclase: Complex with Adenosine-5'-Triphosphate and Ca 3et6 
The crystal structure of the catalytic domain of a eukaryotic guanylate cyclase 3g82 
Complex of GS-alpha with the catalytic domains of mammalian adenylyl cyclase: complex with MANT-ITP and Mn 3maa 
Complex of GS-Alpha with the Catalytic Domains of Mammalian Adenylyl Cyclase: Complex with Adenosine 5-O-(l-Thiophosphate) and Low Ca Concentration 3mr7 
Crystal Structure of Adenylate/Guanylate Cyclase/Hydrolase from Silicibacter pomeroyi 3r5g 
Crystal structure of the adenylyl cyclase CyaB from P. aeruginosa 3uvj 
Crystal structure of the catalytic domain of the heterodimeric human soluble guanylate cyclase 1. 4clf 
Crystal structure of human soluble Adenylyl Cyclase (Apo form) 4clk 
Crystal structure of human soluble Adenylyl Cyclase in complex with alpha,beta-methyleneadenosine-5'-triphosphate 4cll 
Crystal structure of human soluble Adenylyl Cyclase in complex with bicarbonate 4clp 
Crystal structure of human soluble Adenylyl Cyclase complex with adenosine-3',5'-cyclic-monophosphate 4cls 
Crystal structure of human soluble Adenylyl Cyclase with Pyrophosphate 4clt 
Crystal structure of human soluble Adenylyl Cyclase with adenosine-3', 5'-cyclic-monophosphate and pyrophosphate 4clu 
Crystal structure of human soluble Adenylyl Cyclase with pyrophosphate 4clw 
Crystal structure of human soluble Adenylyl Cyclase in complex with alpha,beta-methyleneadenosine-5'-triphosphate soaked with bisulfite 4cly 
Crystal structure of human soluble Adenylyl Cyclase soaked with biselenite 4clz 
Crystal structure of human soluble Adenylyl Cyclase with Inhibitor 4, 4'-Diisothiocyano-2,2'-stilbenedisulfonic acid 4cm0 
Crystal structure of human soluble Adenylyl Cyclase with alpha,beta- methyleneadenosine-5'-triphosphate soaked with bicarbonate 4cm2 
Crystal structure of human soluble Adenylyl Cyclase soaked with bisulfite 4ni2 
Crystal structure of the heterodimeric catalytic domain of wild-type human soluble guanylate cyclase 4oya 
Human solAC Complexed with (4-Aminofurazan-3-yl)-[3-(1H-benzoimidazol-2-ylmethoxy)phenyl]methanone 4oyb 
Crystal Structure Analysis of the solAC 4oyi 
Human solAC Complexed with (4-Amino-furazan-3-yl)-phenyl-methanone 4oym 
Human solAC Complexed with (4-Amino-furazan-3-yl)-(3-methoxy-phenyl)-methanone 4oyo 
Human solAC Complexed with 4-(2-Chlorophenyl)-3-methyl-1H-pyrazole 4oyp 
Human solAC Complexed with 1-Benzofuran-2-carboxylic acid 4oyw 
Crystal Structure of Human Soluble Adenylate Cyclase 4oyx 
Human solAC Complexed with AMPCPP 4oyz 
Human solAC Complexed with Bicarbonate 4oz2 
Human solAC Complexed with 4-(4-Fluorophenyl)-3-methyl-1H-pyrazole 4oz3 
Human solAC Complexed with 4-phenyl-3-(trifluoromethyl)-1H-pyrazole 4p2f 
4P2F 4p2m 
4P2M 4p2x 
4P2X 4ust 
4UST 4usu 
4USU 4usv 
4USV 4usw 
4USW 4wp3 
4WP3 4wp8 
4WP8 4wp9 
4WP9 4wpa 
4WPA 4yus 
4YUS 4yut 
4YUT 5d0e 
5D0E 5d0g 
5D0G 5d0h 
5D0H 5d0r 
5D0R 5d15 
5D15 5iv3 
5IV3 5iv4 
5IV4 - Links (links to other resources describing this domain)
-
PFAM guanylate_cyc INTERPRO IPR001054